Lipide und Vitamine in Chloroplasten höherer Pflanzen: Essentiell für Photosynthese und menschliche Ernährung
Forschungsbericht (importiert) 2004 - Max-Planck-Institut für Molekulare Pflanzenphysiologie
Chloroplasten der Pflanzen enthalten einen einzigartigen Satz von Lipiden
Die Photosynthese der Pflanzen ist die Grundlage allen Lebens auf der Erde, denn Pflanzen sind in der Lage, Lichtenergie in chemische Energie umzuwandeln, die zum Aufbau organisch chemischer Verbindungen notwendig ist. Darüber hinaus wird bei der oxygenen Photosynthese in Pflanzen elementarer Sauerstoff freigesetzt, der für die Atmung der eukaryotischen Organismen unabdingbar ist. Die Chloroplasten höherer Pflanzen enthalten ein einzigartiges Membransystem, die Thylakoide, welche die Chlorophyll-Protein-Komplexe der Photosynthese beherbergen. Die Thylakoid-Membranen unterscheiden sich in ihrer Protein- und Lipid-Zusammensetzung von allen anderen pflanzlichen Membranen, aber auch von den Membranen anderer Organismen wie Bakterien, Hefen oder Tieren. Der Hauptanteil der Thylakoidlipide wird von den Galactolipiden gestellt, die ein oder zwei Galaktose-Zucker gebunden an Diacylglycerol enthalten (Abb. 1).
 und Digalactosyldiacylglyceol (DGDG), stellen die häufigste Lipidklasse der Biosphäre dar. Das Sulfolipid Sulfochinovosyldiacylglycerol (SQDG) enthält einen sulfonylierten Zucker als Kopfgruppe. Phosphatidylglycerol (PG) stellt das einzige Phospholipid der Thylakoide dar. β-Carotin (Provitamin A) gehört zur Lipidklasse der Carotinoide, die in Pflanzen an der Photosynthese beteiligt sind. Zu den Isoprenoidlipiden gehören die Elektronenträger der Photosynthese, Plastochinon und Phyllochinon (Vitamin K) sowie das Antioxidationsmittel Tocopherol (Vitamin E).")
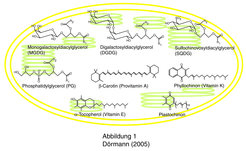
Zusammen machen die Galactolipide circa 75 % der Thylakoidlipide aus. Da die Thylakoide mit Abstand den größten Anteil aller Membranen in den grünen Geweben ausmachen, stellen diese beiden Lipide auch die größte Lipidklasse in Pflanzen und somit in der gesamten Biosphäre dar. Phospholipide dagegen, wie sie aus Hefen und Tieren bekannt sind, kommen in Chloroplasten nur in geringer Menge vor. Das einzige Phospholipid der Thylakoide, Phosphatidylglycerol, macht nur etwa 10 % der Membranlipide aus (Abb. 1). Weiterhin enthalten Chloroplasten circa 5 % Sulfochinovosyldiacylglycerol, ein Sulfoglycolipid, das ausschließlich in Chloroplasten-Membranen vorkommt.
Die Thylakoid-Membranen enthalten weiterhin verschiedene Isoprenoidlipide. Hierzu gehören die grünen Blattpigmente (Chlorophylle) und Carotinoide sowie andere Isoprenoidlipide, die als Elektronenträger der Photosynthese (Plastochinon und Phyllochinon) und als Antioxidationsmittel (Tocopherol) bekannt sind (Abb. 1). Einige der Isoprenoidlipide sind essentiell für die menschliche Ernährung, denn sie stellen Vitamine dar und können nicht vom menschlichen Stoffwechsel synthetisiert werden. β-Carotin, ein Carotinoid der Photosynthesekomplexe, dient als Provitamin A der Erzeugung von Retinol, das als Sehpigment der Augen bekannt ist. Phyllochinon (Vitamin K) dient im Menschen als Kofaktor bei der γ-Carboxylierung von Glutamat-Einheiten, insbesondere in Proteinfaktoren der Blutgerinnung (z.B. Prothrombin). Tocopherol schließlich ist ein hochwirksames Antioxidationsmittel, das die Zellen vor reaktivem Sauerstoff schützt.
Galactolipide sind essentiell für die Photosynthese und für das Wachstum unter Phosphatmangel
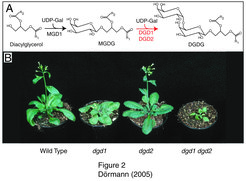
Die beiden Galactolipide Monogalactosyldiacylglycerol (MGDG) und Digalactosyldiacylglycerol (DGDG) bilden das Rückgrat der Thylakoidmembran (Abb. 1); [1]. Außerdem kommen sie gebunden an die Photosynthesekomplexe vor. So wurde MGDG als Bestandteil der Struktur von Photosystem I gefunden. DGDG ist ein integraler Bestandteil der Lichtsammelkomplexe II (light harvesting complexes, LHC II). Die große Menge an Galactolipiden in den Thylakoid-Membranen sowie ihr Vorkommen in den Photosynthesekomplexen ließ schon früh vermuten, dass sie für die photosynthetischen Lichtreaktionen essentiell sind. Eine Bestätigung dieser Hypothese erhielten die Wissenschaftler aus der Untersuchung von Galactolipid-defizienten Arabidopsis-Mutanten. Die mgd1-Mutante von Arabidopsis enthält nur noch etwa die Hälfte des normalen MGDG-Gehalts [2]. Die Pflanzen sind im Wachstum und in der Photosynthese-Effizienz reduziert und enthalten weniger Chlorophyll. Dies zeigt, dass MGDG essentiell für die Aufrechterhaltung der normalen Photosynthese ist. In Arabidopsis gibt es zwei Gene, die für Enzyme der DGDG-Synthese kodieren: DGD1 und DGD2 [3]. DGD1 ist für die Synthese des größten Teils an DGDG verantwortlich. Dementsprechend enthält die dgd1-Mutante nur noch 1/10 des normalen DGDG-Gehalts und ist im Wachstum und in der Photosynthese-Effizienz beeinträchtigt (Abb. 2). Die Doppelmutante, dgd1 dgd2 ist völlig DGDG-frei und dramatisch in der Photosynthese-Effizienz beeinträchtigt. Ein deutliches Indiz dafür, dass das Galactolipid DGDG für die Photosynthese essentiell ist [4].
 Monogalactosyldiacylglycerol (MGDG) wird aus Diacylglycerol und UDP-Galactose durch MGDG-Synthasen (z. B. MGD1) synthetisiert. Die beiden DGDG-Synthasen, DGD1 und DGD2, katalysieren die Umsetzung von MGDG zu Digalactosyldiacylglycerol (DGDG). B) Die Arabidopsis-Mutanten dgd1, dgd2 und die Doppelmutante dgd1 dgd2 zeigen verschiedene Grade an Wachstumsreduktion.")
Neben der Photosynthese spielen Galactolipide eine wichtige Rolle beim Wachstum von Pflanzen unter Phosphatmangel. Pflanzen benötigen große Mengen an Phosphat zur Aufrechterhaltung ihres Stoffwechsels. Ein erheblicher Teil des Phosphats wird für die Synthese der Nukleinsäuren RNA und DNA verwendet, also für die Genexpression und die Replikation des Genoms. Weiterhin findet sich organisch gebundenes Phosphat in Proteinen und in den Nucleotiden (z. B. ATP, GTP), phosphorylierten Zuckern und Zucker-Nucleotiden (z. B. UDP-Glucose). Ein großer Anteil des Phosphats wird aber für die Synthese von Phospholipiden, also für die Ausbildung der biologischen Membran verwendet. Unter Phosphatmangel werden in Pflanzen Phospholipide in den Membranen abgebaut und durch phosphatfreie Glycolipide (DGDG und SQDG) ersetzt. Das Phosphat aus den Membranen wird für andere wichtige Prozesse verwendet, beispielsweise zur RNA- und DNA-Synthese. Die Umwandlung der Phospholipide in Glycolipide stellt einen aktiven Prozess dar, weil die Expression einer Reihe von Genen der Galactolipid- und der Sulfolipid-Biosynthese induziert wird. Dieser elementare Prozess des Austausches von Phospholipiden und Glycolipiden betrifft nicht nur die chloroplastidären Membranen, sondern auch extraplastidäre Membranen wie die Plasmamembran oder Mitochondrien.
Vitamin-E-Mangel in Pflanzen hat nur einen geringen Einfluss auf Wachstum und Photosynthese
Die Gruppe der Tocopherole und Tocotrienole bildet die Stoffklasse des Vitamin E, das als Antioxidationsmittel in Pflanzen und Tieren eine große Bedeutung beim Abbau von reaktivem Sauerstoff (reactive oxygen species) wie Peroxiden, Superoxiden und Singulett-Sauerstoff spielt. Die Kopfgruppe des Tocopherols stammt aus dem Biosyntheseweg der aromatischen Aminosäuren („Shikimat“-Biosyntheseweg), während die Seitenkette aus der Isoprenoid-Synthese abgeleitet ist (Abb. 3); [5].
-Mangel hat keinen Einfluss auf das Wachstum von Arabidopsis-Pflanzen.
A) Biosynthese von α-Tocopherol aus p-Hydoxyphenylpyruvat. B) Eine Mutation im Gen der Tocopherol-Cyclase (VTE1) führt zum vollständigen Verlust von Tocopherol in der Arabidopsis-Mutante vte1, hat aber keinen großen Einfluss auf das Wachstum.")
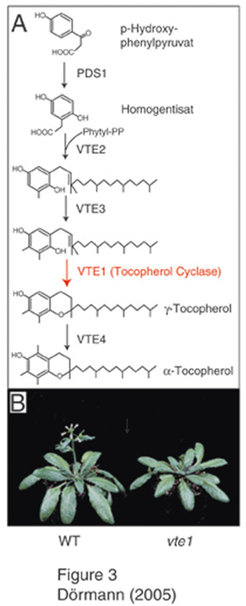
Je nachdem ob der Isoprenoidteil von Phytol oder von Geranylgeranol abgeleitet ist, unterscheidet man Tocopherole mit gesättigter Seitenkette bzw. Tocotrienole mit ungesättigter Seitenkette [6]. In Pflanzen wird Tocopherol in den Hüllmembranen der Chloroplasten synthetisiert. Es schützt ungesättigte Fettsäuren und die Chlorophyll-Protein-Komplexe der Photosynthese in den Thylakoiden vor Oxidation. Durch einen genetischen Screeningansatz wurden Arabidopsis-Mutanten isoliert, in denen die Tocopherol-Biosynthese gestört ist. Die vte1-Mutante (für Vitamin-E-defizient 1), die im Rahmen dieses Projekts gefunden wurde, enthält kein Tocopherol mehr und stellt somit die erste Vitamin-E-freie Pflanze dar [7]. Erstaunlicherweise sind das Wachstum, der Gehalt an ungesättigten Fettsäuren und die Photosynthese-Effizienz in diesen Pflanzen kaum verändert. Daher muss geschlussfolgert werden, dass Tocopherol für die Photosynthese der Pflanze nicht absolut notwendig ist. Es gibt offensichtlich alternative Antioxidationsmittel, die an der Abwehr von reaktivem Sauerstoff beteiligt sind (zum Beispiel Glutathion, Ascorbat, Carotinoide; [8]). Im Gegensatz zu grünen Blättern ist Tocopherol allerdings bei der Samenkeimung essentiell. Untersuchungen zeigten, dass Tocopherol-Mangel die Keimungsfähigkeit der Samen dramatisch verringert, wahrscheinlich durch eine vermehrte Oxidation des Speicherlipids durch reaktiven Sauerstoff [9].
Das Gen, welches in der vte1-Mutante betroffen ist, wurde im Arabidopsis-Genom kartiert und durch Komplementation isoliert [7]. Funktionstests ergaben, dass es sich bei dem VTE1-Protein um die Tocopherol-Cyclase handelt, die für den vorletzten Schritt der Tocopherol-Biosynthese in Pflanzen wichtig ist. Die Tocopherol-Cyclase katalysiert den Schluss des zweiten Rings des Tocopherol-Moleküls und ist daher für die Synthese der charakteristischen Kopfgruppe notwendig. Weiterhin ist die Tocopherol-Cyclase maßgeblich an der Regulation der Tocopherol-Biosynthese beteiligt. Die Überexpression von VTE1 in transgenen Arabidopsis-Pflanzen ergab eine Steigerung des Tocopherol-Gehalts in den Blättern [8]. Die Gene der Tocopherol-Cyclase und anderer Schritte der Tocopherol- und Tocotrienol-Biosynthese werden zurzeit für die Erhöhung des Vitamin-E-Gehalts in transgenen Nutzpflanzen verwendet, um den Bedarf dieses wichtigen Antioxidants für die menschliche Ernährung in den Industriestaaten und in der dritten Welt sicherzustellen (z. B. [6]; [10]). Ein großer Teil des Vitamin E, das als Nahrungsergänzungsstoff im Handel ist, wird synthetisch aus Erdölprodukten gewonnen und unterscheidet sich in seiner Isomeren-Zusammensetzung von natürlich vorkommendem Vitamin E. Mithilfe biotechnologischer Methoden sollte es aber in Zukunft möglich sein, den Bedarf an natürlichem Vitamin E aus transgenen Nutzpflanzen zu decken.