Gene-Silencing in transgenen Pflanzen
Forschungsbericht (importiert) 2005 - Max-Planck-Institut für Molekulare Pflanzenphysiologie
Mechanismen der Genregulation beruhen oft auf Wechselwirkungen von Proteinen mit bestimmten DNS-Sequenzen, jedoch sind auch Mechanismen der Inaktivierung für Gene beschrieben worden, die nicht direkt durch eine bestimmte Basenabfolge der DNS-Sequenz vermittelt werden. Es handelt sich dabei um so genannte epigenetische Regulationsmechanismen. Diese Phänomene werden im Pflanzenreich seit vielen Jahren intensiv bearbeitet. So wird beispielsweise bei der Untersuchung transgener Pflanzen häufig die Inaktivierung (Silencing) eingeführter Gene beobachtet. Es ist von großem Interesse, die verantwortlichen Mechanismen aufzuklären, da diese Inaktivierung die Nutzung von transgenen Pflanzen in Forschung und Biotechnologie limitiert.
Ein Untersuchungssystem zum Silencing in transgenen Pflanzen
Der Aufbau eines Testsystems in Arabidopsis thaliana (Ackerschmalwand) ermöglichte es, die Ausprägung verschiedener Transgene in unterschiedlicher, aber definierter Anzahl in verschiedenen Positionen des Genoms der Modellpflanze zu untersuchen. Als Transgene wählten die Nachwuchsforscher am Max-Planck-Institut für molekulare Pflanzenphysiologie Gene aus, die für die Glucuronidase (GUS) oder das grün-fluoreszierende Protein (GFP) kodieren. Für diese Gene kann durch einfache Farb- beziehungsweise Fluoreszenznachweise sehr genau bestimmt werden, wie intensiv sie in den verschiedenen Teilen einer Pflanze ausgeprägt werden. Die Transgene wurden unter die Kontrolle eines Regulationselementes, den Promotor gestellt, der eine hohe Expression des eingeführten Gens in allen Teilen der Pflanze vermittelt. Nach Übertragung der Transgene in das Erbmaterial von Arabidopsispflanzen führten die Wissenschaftler eingehende genetische und molekulargenetische Untersuchungen durch, um diejenigen Pflanzen auszuwählen, welche jeweils nur eine intakte Kopie eines bestimmten Transgens im Genom trugen. Dies ist nötig, da es gängige Genübertragungsmethoden nicht ermöglichen, eine definierte Anzahl von Transgenkopien in das Erbmaterial einer höheren Pflanze einzubringen. Die Integration an einer vorbestimmten Stelle ist ebenfalls nicht effizient zu gewährleisten. Daher unterscheiden sich verschiedene transgene Pflanzen nicht nur bezüglich der Anzahl der eingeführten Transgene, sondern auch durch die Position, an der die Transgene im Erbmaterial eingelagert sind. Für die ausgewählten transgenen Pflanzen, die nur eine Kopie eines eingeführten Gens trugen, wurde die exakte Position im Erbmaterial ermittelt. Durch Kreuzungen verschiedener transgener Linien konnten Pflanzen erzeugt werden, die sich hinsichtlich der Kopienzahl im Genom unterschieden.
Übermäßige Ausprägung führt zur Inaktivierung des Transgens
Die Untersuchungen zeigten, dass verschiedene transgene Linien mit entweder einer Kopie des GUS- oder des GFP-Transgens eine hohe Expression des eingeführten Gens aufwiesen. Unabhängig erstellte transgene Linien unterschieden sich nur wenig hinsichtlich der Stärke der Genausprägung. Da sich die verschiedenen untersuchten Linien hinsichtlich der Integrationsstelle des Transgens im Erbmaterial unterschieden, spielt demnach die Genomposition in der Regel eine untergeordnete Rolle bezüglich der Expression eines eingeführten Transgens, In all jenen Nachkommen, die genau wie die Elternpflanzen nur eine Kopie des Transgens aufwiesen, beobachteten die Forscher eine ähnlich hohe Ausprägung des Transgens.
Im Gegensatz dazu können sich unterschiedliche Kopienzahlen eines Transgens deutlich auf die Genexpression auswirken. Interessanterweise zeigen sich sowohl positive als auch negative Korrelationen zwischen der Kopienzahl eines Transgens und seiner Ausprägung. Unterhalb eines bestimmten Schwellenwertes ist die Ausprägung eines Transgens direkt proportional zur Kopienzahl, während das Überschreiten des Schwellenwertes die gleichzeitige Inaktivierung aller Transgenkopien auslöst. Untersucht man die Transkriptspiegel in Pflanzen, die Silencing der eingeführten Gene zeigen, so findet man, dass hier nur noch sehr wenige Transkripte des Transgens nachweisbar sind (Abb. 1).
, die drei oder vier Kopien des Gens für das grün-fluoreszierende Protein (GFP) unter der Kontrolle eines Promotors trugen, der hohe Expression in allen Pflanzenteilen vermittelt. Unter (b) sind Untersuchungen der Transkriptspiegel zu sehen. Der Grad der Schwärzung spiegelt dabei die Höhe der Transgenexpression wider. Durch Einbringen weiterer Genkopien wurde die Inaktivierung der Transgene ausgelöst. In diesen Pflanzen waren nur wenige GFP-Transkripte zu sehen (b) und grüne Fluoreszenz war nicht nachweisbar. Die rote Fluoreszenz, die in diesen Pflanzen zu erkennen ist, wird durch Chlorophyll hervorgerufen (a).")
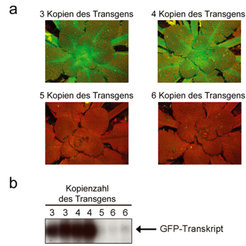
Um den beobachteten Schwellenwert der Inaktivierung von Transgenen näher zu untersuchen, etablierte das Team um Renate Schmidt Pflanzen mit gleicher Kopienzahl der Transgene, die sich jedoch in Bezug auf den Promotor unterschieden. Die ausgewählten Promotoren vermittelten verschieden hohe Ausprägung der Transgene. Diese Experimente zeigten eindeutig, dass nicht die Anzahl der eingeführten Transgene darüber entscheidet, ob die Gene stillgelegt werden, sondern die Höhe der Expression, die durch die verschiedenen Kopien vermittelt wird. Die Ausprägung übermäßiger Transkriptmengen in Pflanzenzellen führt zur Überschreitung des Schwellenwertes und somit zur Auslösung von Transgene-Silencing.
Ausbreitung von Silencing
Je höher die Ausprägung der eingeführten Transgene, desto größer ist die Wahrscheinlichkeit, dass Silencing früh in der pflanzlichen Entwicklung ausgelöst wird. Diese Inaktivierung oder Stilllegung von Genen kann besonders gut in Pflanzen beobachtet werden, deren Transgene das grün-fluoreszierende Protein ausprägen. Mithilfe eines Fluoreszenzstereomikroskopes können Bereiche einer Pflanze mit GFP-Fluoreszenz leicht von denen unterschieden werden, die keine GFP-Fluoreszenz aufweisen. Solche Untersuchungen machten sichtbar, dass Silencing nicht gleichzeitig in allen Zellen einer Pflanze, sondern lokal ausgelöst wird. Selbst in genetisch identischen Individuen variiert sowohl der Zeitpunkt als auch der Teil der Pflanze, in dem die Inaktivierung einsetzt. Ist der Prozess einmal in Gang, so breitet er sich von Zelle zu Zelle und über die Leitgewebe weiter aus, sodass es nach Tagen oder Wochen zum vollständigen Verlust der Transgenexpression in der Pflanze kommen kann (Abb. 2).
 die Transgeninaktivierung auf, so beginnt dies als lokales Ereignis (b), wobei Ort und Zeitpunkt der Inaktivierung selbst bei genetisch identischen Pflanzen unterschiedlich sind. Silencing breitet sich mit der Zeit aus (c, d), sodass ganze Blätter (e) und schließlich die gesamte Pflanze betroffen sind. Dieser Prozess kann Tage bis Wochen dauern.")

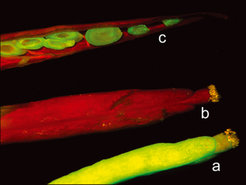
Die Untersuchung von Nachkommen der Pflanzen, die Silencing aufwiesen, ergab, dass der inaktivierte Zustand der Transgenkopien nicht vererbt wird (Abb. 3). Erst im Laufe der Entwicklung der Nachkommenpflanzen wird Transgene-Silencing erneut etabliert.
 und (c) gezeigten Schoten transgener Arabidopsispflanzen wiesen Inaktivierung der GFP-Transgene auf, die Samen in der geöffneten Schote (c) zeigten jedoch GFP-Fluoreszenz und somit die Reaktivierung der GFP-Transgene in den Nachkommen. Zum Vergleich ist unter (a) die Schote einer Pflanze mit hoher und stabiler GFP-Expression gezeigt.")
Mechanismus der Geninaktivierung
Epigenetische Regulationsmechanismen werden intensiv in transgenen Pflanzen bearbeitet, sind jedoch keine Besonderheit von Transgenen, sondern kommen im gesamten Pflanzenreich und vielen weiteren Organismen vor. So konnten ausgeprägte Parallelen zwischen der Inaktivierung von Transgenen und einigen Formen der Virusabwehr in Pflanzen sowie der entwicklungsspezifischen Regulation pflanzeneigener Gene gefunden werden. Außerdem wurde festgestellt, dass Silencing-Prozessen in Pilzen, Pflanzen und Tieren sehr ähnliche Mechanismen zu Grunde liegen. Doppelsträngige RNS-Moleküle lösen Silencing auf effiziente Art und Weise aus. So wurde für transkribierte invertierte Sequenzwiederholungen nachgewiesen, dass die gebildete doppelsträngige RNS zu kurzen RNS-Molekülen einer bestimmten Länge, so genannter kleiner interferierender RNS, abgebaut wird. Diese kleinen RNS-Moleküle bewirken Gene-Silencing durch den sofortigen Abbau von Transkripten mit komplementären Basenabfolgen. Sie können aber auch eine Modifikation von DNS-Sequenzen hervorrufen, die Sequenzähnlichkeit zu den kleinen interferierenden RNS-Molekülen aufweisen. Die Modifikation besteht darin, dass ausgewählte Cytosinbasen mit einer Methylgruppe versehen werden.
Die beschriebenen Ergebnisse zum Transgene-Silencing in Arabidopsis zeigen eindeutig, dass es einen weiteren Weg gibt, durch den die Inaktivierung von Genen ausgelöst werden kann. Nicht nur die Bildung doppelsträngiger RNS und ihre Prozessierung führt zur Stilllegung, sondern auch die übermäßige Transkription von Transgenen. Molekularbiologische Untersuchungen zeigen, dass beiden Wegen ähnliche molekulare Mechanismen zu Grunde liegen. Wird Silencing durch übermäßige Transkriptspiegel ausgelöst, findet man kleine interferierende RNS-Moleküle. Diese sind komplementär zu den transkribierten Transgensequenzen und bewirken den sofortigen Abbau der gebildeten Transgentranskripte sowie die Methylierung von einigen Cytosinbasen im transkribierten Bereich der Transgene. In Pflanzen mit hoher und stabiler Transgenexpression konnten dagegen weder kleine interferierende RNS-Moleküle noch Methylierung der Transgensequenzen nachgewiesen werden.
Wie die übermäßige Transkriptmenge in Zellen wahrgenommen wird und zur Auslösung von Silencing führt, ist bisher nicht geklärt. Die Wissenschaflter am MPI in Potsdam konnten aber zeigen, dass die Schwellenwerte, bei denen die Inaktivierung ausgelöst wird, spezifisch für die jeweils untersuchten Gene sind. Während drei Kopien eines GUS-Transgens unter der Kontrolle eines starken Promotors des Blumenkohlmosaikvirus ausreichen, um die Stilllegung aller Transgene in Arabidopsis zu bewirken, wird der gleiche Effekt erst durch die Einführung von fünf GFP-Transgenen unter der Kontrolle desselben Promotors erzielt. Die Aufklärung der Faktoren, welche die genspezifischen Schwellenwerte bestimmen, ist ein wichtiges Ziel künftiger Untersuchungen, da dies unabdingbar ist, um Strategien zur Vermeidung oder Verhinderung des Transgene-Silencing zu entwickeln. Solche Strategien würden für die Nutzung transgener Pflanzen in der Biotechnologie einen großen Fortschritt darstellen. Aber auch für die Grundlagenforschung sind diese Erkenntnisse von großem Wert, da transgene Pflanzen zur Aufklärung von Genfunktionen herangezogen werden.
Einfluss von Umweltfaktoren auf die Transgeninaktivierung
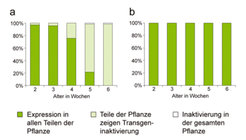
Pflanzen als sesshafte Organismen haben sehr effektive Strategien entwickelt, um unter wechselnden Umweltbedingungen überleben zu können. Es häufen sich die Anzeichen, dass auch epigenetischen Regulationsphänomenen hierbei eine wichtige Rolle zukommt. Mit Hilfe des vorgestellten Testsystems können die Wechselwirkungen von Umweltbedingungen und epigenetischer Regulation erstmals systematisch erforscht werden. So konnte bereits gezeigt werden, dass in Pflanzen, die in der Kälte angezogen wurden, die Auslösung von Silencing sehr stark reduziert und verzögert ist (Abb. 4). Die beschriebenen Ergebnisse verdeutlichen, welch wichtige Rolle die Aufklärung von Silencingprozessen in der Pflanzenforschung, Biotechnologie und Landwirtschaft in Zukunft haben kann.
 als Pflanzen, die bei 4 °C angezogen wurden (b). Jeweils 70 Pflanzen wurden in wöchentlichen Abständen untersucht und die Balkendiagramme geben den Anteil der Pflanzen an, welche die angegebenen Ausprägungsmuster aufwiesen.")