Rekonstruktion der Genom-Evolution im Labor
Forschungsbericht (importiert) 2006 - Max-Planck-Institut für Molekulare Pflanzenphysiologie
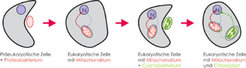
Pflanzenzellen besitzen insgesamt drei Genome: ein großes im Zellkern und zwei deutlich kleinere in den Mitochondrien und Plastiden (Chloroplasten). Mitochondrien und Plastiden haben eine faszinierende Entstehungsgeschichte. Sie stammen von ursprünglich selbständigen Lebewesen ab, die vermutlich vor über einer Milliarde Jahren von einer so genannten präeukaryotischen (eukaryotisch = mit Zellkern) Vorläuferzelle verschluckt, aber nicht verdaut wurden: Zunächst wurde ein Proteobakterium von einer Einstülpung der Zellmembran umschlossen und schließlich vollständig in das Innere, das Protoplasma der präeukaryotischen Wirtszelle aufgenommen. Die Wirtszelle hat das verschluckte Proteobakterium dann kompromisslos versklavt: Das Bakterium verlor seine Unabhängigkeit, wurde zum Mitochondrium umfunktioniert und diente fortan seiner Wirtszelle zur Energiegewinnung durch Atmung. Diesen Zelltyp mit zwei Genomen (im Zellkern und in den Mitochondrien) finden wir heute in allen Tieren und Pilzen. Bei der Entstehung der Pflanzenzelle wiederholte sich die Versklavung eines Bakteriums, die auch als Endosymbiose bezeichnet wird, ein weiteres Mal (Abb.1). Nun wurde ein Cyanobakterium (eine Blaualge, die bereits zur Photosynthese befähigt war, also Lichtenergie in chemische Energie umwandeln konnte) aufgenommen und zum zweiten Typ DNA-haltiger Zellorganellen, den Plastiden, umfunktioniert. Plastiden und Mitochondrien sind aufgrund dieser Entstehungsgeschichte von zwei Membranen umschlossen: die äußere stammt von der Wirtszelle, die innere vom aufgenommenen Bakterium.
 und Chloroplasten (grün)
Vorläufer der Organellen waren freilebende Bakterien, die von einer „Urzelle“ (=präeukaryotische Zelle) umschlossen wurden. Auf diese Weise sind aus so genannten Proteobakterien die für die Atmung zuständigen Mitochondrien entstanden und aus Cyanobakterien (Blaualgen) die für die Photosynthese verantwortlichen Chloroplasten. Die Pfeile innerhalb der Zellen verdeutlichen Richtung und Umfang des Gentransfers zwischen den Genomen der Pflanzenzelle: Riesige Mengen genetischer Information wurden im Verlauf der Evolution aus den Plastiden- und Mitochondriengenomen in das Kerngenom übertragen, kleinere Mengen an Information auch aus den Plastiden in die Mitochondrien sowie aus dem Zellkern in die Mitochondrien.")
Nach der Endosymbiose erfolgte im Zuge der allmählichen Umwandlung der aufgenommenen Bakterien in Zellorganellen eine hochgradig komplexe genetische Umstrukturierung aller beteiligten Genome. Eine gewaltige Menge an bakteriellen (prokaryotischen) Genen wurde im Verlauf der Evolution aus den Genomen der aufgenommenen Bakterien in das Kerngenom der Wirtszelle transferiert (Abb. 1). Dies hat dazu geführt, dass die Organellengenome heute nur noch einige Dutzend Gene enthalten, obwohl die Bakterien, aus denen sie hervorgegangen sind, vermutlich mindestens ein paar Tausend Gene besaßen [1, 2]. Indirekte Hinweise auf diesen Gentransfer aus den Mitochondrien- und Plastidengenomen in das Kerngenom haben Sequenzähnlichkeiten zwischen Kerngenen und Genen aus Proteobakterien bzw. Cyanobakterien geliefert. Auch gibt es einige Fälle von offensichtlich erst vor relativ kurzer Zeit transferierten Genen. Solche Gene liegen bei einigen Pflanzenarten noch in den Organellen vor, während sie bei anderen Arten aber bereits im Kerngenom zu finden sind [2].
Wie kann ein Gen aus dem von zwei Membranen umschlossenen Plastiden- oder Mitochondriengenom in das Kerngenom gelangen? Da die Übertragung tausender Gene aus den Organellen in den Kern offenbar in riesigen evolutionären Zeiträumen ablief und demzufolge niemand jemals ein solches Ereignis beobachten konnte, entzog sich diese Frage bislang einem experimentellen Zugang. Die Entwicklung neuer Technologien, die es erlauben, Plastidengenome höherer Pflanzen gentechnisch zu verändern [3, 4, 5], hat es in den letzten Jahren ermöglicht, wichtige Schritte dieses evolutionären Prozesses im Labor nachzuvollziehen und die molekularen Grundlagen des Gentransfers zwischen Organellen- und Kerngenomen zu analysieren.
Gentransfer im Zeitraffer
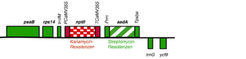
In einem Schlüsselexperiment zur Rekonstruktion des Gentransfers im Labor wurden Chloroplastengenome in der Modellpflanze Tabak gentechnisch verändert, indem zwei zusätzliche Gene eingebracht wurden. Während eins der beiden Gene, aadA genannt, für das Chloroplastengenom maßgeschneidert ist durch seine bakterielle (prokaryotische) Genstruktur und eine Resistenz gegen das Antibiotikum Spectinomycin vermittelt, ist das zweite Gen, nptII, für den Zellkern konstruiert worden (eukaryotische Genstruktur) und verleiht den Pflanzen, im Kern platziert, eine Resistenz gegen das Antibiotikum Kanamycin (Abb.2).
, das plastidäre Expressionssignale besitzt (den Plastidenpromotor Prrn und den Plastidenterminator PpsbA) sowie ein Kanamycin-Resistenzgen (nptII), das über nukleäre Expressionssignale verfügt (Promotor und Terminator des 35S-Gens aus dem Pflanzenvirus CaMV). psaB, rps14, trnG und ycf9 sind benachbarte Gene im Plastidengenom.")
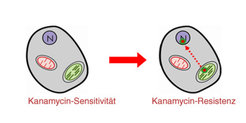
Im Chloroplasten ist nptII dagegen nicht aktiv. Pflanzen mit transgenen Plastidengenomen sind demzufolge zwar resistent gegen Spectinomycin, reagieren aber empfindlich auf Kanamycin. Wenn Zellen dieser Pflanzen in einem Gewebekultursystem einer Selektion auf Kanamycinresistenz ausgesetzt werden, so sollten nur solche Zelle überleben können, die das nptII-Gen aus dem Plastidengenom in das Kerngenom verschoben haben (Abb.3).

Mit dieser experimentellen Strategie gelang es in der Tat, Pflanzen zu selektieren, die das nptII-Gen in den Zellkern transferiert hatten [6]. Genetische und molekulare Untersuchungen bestätigten, dass das Gen im Kerngenom angekommen war: Die Kanamycinresistenz wurde nach den Mendel'schen Regeln vererbt (während Chloroplastengene mütterlich vererbt werden) und die DNA des nptII-Gens konnte im Genom des Zellkerns nachgewiesen werden. Erstaunlich hoch war die Frequenz, mit der ein solcher Gentransfer aus dem Chloroplasten in den Zellkern detektiert wurde: In ungefähr einer von 5 Millionen Zellen war das nptII-Gen in den Zellkern gesprungen [6] (Abb.4).
 oder als regenerierende Sprosse (rechts). Das umgebende bleiche Gewebe stirbt aufgrund seiner Empfindlichkeit gegen Kanamycin ab.")

Worin bestehen die Konsequenzen einer solch hohen Rate an Gentransfer zwischen den Organellen und dem Zellkern? Zum einen verändern die Befunde unser Verständnis von genetischer Homogenität innerhalb einer Art und innerhalb eines Organismus. Dies wird offensichtlich, wenn man sich klarmacht, dass zum Beispiel ein Tabakblatt aus wesentlich mehr als fünf Millionen Zellen besteht. Die Zellen in ein und demselben Blatt einer Pflanze sind folglich nicht notwendigerweise genetisch identisch, sondern können sich im Muster der in das Kerngenom transferierten Organellensequenzen unterscheiden. Eine weitere interessante Konsequenz ergibt sich aus dem zufälligen Einbauort transferierter Organellengene im Kerngenom. Mit einer gewissen Wahrscheinlichkeit kann der Einbau in ein Gen des Kerngenoms erfolgen und dieses damit zerstören. Somit könnte der Transfer von Organellen-DNA in den Zellkern auch zum spontanen Auftreten von Mutationen (so genannten „somatischen Mutationen“) beitragen.
Wie werden transferierte Gene aktiv?
Die Übertragung eines Gens aus den Plastiden in den Kern führt zunächst nicht zu einem neuen funktionsfähigen Kerngen. Wie am Beispiel der beiden Resistenzgene aadA und nptII erläutert, liegt dies daran, dass sich Plastidengene und Kerngene fundamental in ihrer Struktur unterscheiden: Ein Plastidengen hat eine bakterielle (prokaryotische) Genstruktur und kann daher nicht im Zellkern funktionieren, wo die Gene eine eukaryotische Genstruktur aufweisen. Im oben beschriebenen Schlüsselexperiment wurde dieses Problem umgangen, indem das nptII-Gen mit eukaryotischen Expressionssignalen (Promotor und Terminator; Abb. 2) versehen wurde und so direkt nach seinem Transfer aus dem Plastiden- in das Kerngenom aktiv sein konnte. Beim evolutionären Gentransfer wäre dies nicht der Fall: Das Gen würde im Zellkern ankommen und könnte nicht abgelesen werden, d.h. nicht in eine Boten-RNA übersetzt werden. Wie kann aus einem solchen inaktiven ehemaligen Chloroplastengen ein funktionsfähiges Kerngen werden?
Diese Frage wurde einer experimentellen Analyse zugänglich, als es gelang, Pflanzenlinien zu isolieren, die ein so großes Stück Plastiden-DNA in den Kern übertragen hatten, dass darauf nicht nur das Kanamycin-Resistenzgen nptII, sondern auch das benachbarte Spectinomycin-Resistenzgen aadA lag (Abb. 2). Das aadA-Gen liefert nun genau das geeignete Untersuchungsobjekt: Es ist ein prokaryotisches Gen (mit einem Plastidenpromotor und -terminator) und ist im Zellkern nicht aktiv. Folglich sind die Gentransferpflanzen mit dem aadA-Gen im Zellkern empfindlich gegen das Antibiotikum Spectinomycin. In einem weiteren groß angelegten Selektionsexperiment wurde nun untersucht, ob und wie dieses inaktive aadA-Gen im Kern funktionsfähig werden kann. Dazu wurden Zellen einer Selektion auf spectinomycinhaltigem Kulturmedium ausgesetzt. Auf diese Weise gelang es tatsächlich, Pflanzen zu erhalten, in denen das aadA-Gen im Zellkern aktiv geworden war [7].
Als nächstes galt es nun, die molekularen Mechanismen aufzuklären, die zur Aktivierung des aadA-Gens geführt hatten. Dabei stellte sich heraus, dass in allen acht selektierten Pflanzenlinien sehr ähnliche Vorgänge an der Genaktivierung beteiligt waren. In allen Fällen war es dem aadA-Gen gelungen, durch relativ einfache Mutationen (sog. Deletionen), den Promotor des benachbarten Kerngens „einzufangen“ und nun für seine eigene Expression zu nutzen. Erstaunlicherweise reichte dieser eine molekulare Umbau aus, um das Gen im Zellkern funktionsfähig zu machen. Ähnliche Veränderungen im Terminatorbereich des Gens waren nicht nachweisbar. Tatsächlich ergab eine Analyse der aadA Boten-RNA, dass der Plastidenterminator von der Genexpressionsmaschinerie des Zellkerns benutzt werden konnte, obwohl die molekularen Mechanismen zum Stoppen des Ablesevorgangs im Zellkern völlig anders sind als im Chloroplasten [7].
Diese Experimente haben erste Einsichten in die molekularen Prozesse geliefert, die die Genome der eukaryotischen Zelle seit ihrer Geburt vor mehr als einer Milliarde Jahren geformt haben. Mit neuen Methoden und gentechnischen Verfahren ist es nun möglich, diese Prozesse im Labor in einem Zeitraum von nur wenigen Jahren ablaufen zu lassen. Das eröffnet die aufregende Perspektive, Vorgänge, die normalerweise nur in riesigen evolutionären Zeiträumen ablaufen, im Zeitraffer experimentell nachzuvollziehen und die zugrunde liegenden Mechanismen aufzuklären.